 |
| A normal array of plants in the site: normal purple Trichostema laxum, an individual with the common white and purple lower lip phenotype and some Zeltnera trichantha intermixed (a really cool plant) |
My main question in the system is: how is this polymorphism in flower color maintained? If it were a fitness benefit, we might expect a high proportion of it. If it were deleterious, it should be lost (especially as it is at reasonably low frequency). If it is neutral, it might be drifted out. I actually suspect the answer is quite a bit more complicated.
 |
| Once you start looking for variation, you find it! I don't know what this doubled lower lip is about (it showed up in a plant grown in the greenhouse - on most flowers). The plant was male-sterile, I believe. I'll be looking for it in the field though! |
The first question is, of course, how common is the color polymorph? I censused the focal patch/population (separated by ~300 meters from others) in 2014 and 2015. In 2014, the polymorph was 2.0% (46/2278 individuals) in 2015, 3.7% (102/2757). Neither of these censuses was a complete census of the population - necessarily, I cannot assess the phenotype of any pre- or post-flowering individuals. Both were done roughly in the peak flowering time (over several days), so I do think it is close to accurate. I think its safe to conclude that the proportion stayed the same or even went slightly up in 2015.
 |
| A rather large w/p morph individual. |
The next logical question is: do the two morphs have similar field fitness? Any "fitness" measure (e.g. reproductive success, height, etc.) of these plants is dictated mostly by microhabitat location. In this rocky, heavily serpentine site, most plants stay under 20 cm tall and never put out more than 50 flowers (mints have 4 ovaries per flower, so maximum seed set is four times flower number). In a wetter, less serpentine and rocky meadow, I once found a plant on a gopher mound (which brings up nutrients) that was nearly a meter wide and better than a half meter tall. It probably had >5000 flowers throughout the season.
 |
| A veritable field of Trichostema! Not my field site - this site has huge plants (~500 flowers/plant) and very little flower color variation. It is a nice place to look at the insect communities on T. laxum, as it has really high densities of herbivores and predators (T. laxum gets most of the sticky plant predators) |
Because of this microhabitat variation, the best comparison to make is nearest neighbors which differ in flower color. In both 2014 and 2015, I took data on 41 pairs (coincidentally!) of white/purple and purple/purple neighbors. I found no significant differences, either year, between any fitness variables (number of buds, flowers, fruit, height and, in 2014, number of leaves and herbivory [too low in 2015]).
 |
| A more normal-sized (for this population) individual. |
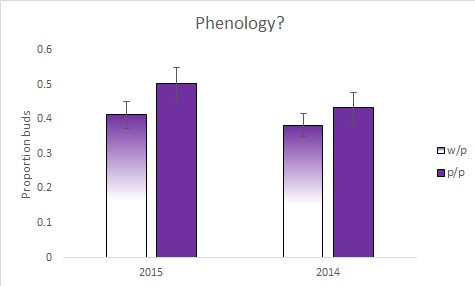
This, ostensibly, seems like the trait is fitness neutral (and lab growouts seem to bear this out - more data soonish). Given that this site burned in August this year (after most had flowered, but some [probably few] were still maturing seeds), I was curious about whether the morphs differed in phenology. Hindsight is 20/20 (I should have censused biweekly!), but the neighbor pairs data can be used to examine this in a roundabout way; I have data on buds, flowers and fruit, so later phenology plants should have a higher proportion of buds to flowers and fruit than earlier phenology plants.
In both years, the white/purple plants had a lower proportion of buds than the purple plants (it is marginally significant). This suggests that they have a earlier phenology - which could be what is under selection - not the flower color itself. I am super, super, super, excited about this (the only positive result so far from anything in T. laxum) - there was possibly a big selective event (a fire) on 12-August (I think - could have burned on the 13th). From this, I'd predict that the w/p morph may have dehisced a higher proportion of their seed set by the fire. I'll be paying far more attention to the phenology, and recensusing more often this upcoming season.
I also analyzed the pollinator data from 2014 and got no particularly useful insights (I wondered if there was some degree of isolation between the morphs). The pollinator communities using each morph were pretty similar and the only real differences were: a bee on a purple flower was more likely to visit a w/p next than a bee starting on a w/p* and, only bees that started on w/p flowers next visited a snapdragon, Antirrhinum cornutum (but only 3% of the time). This last result is interesting as the snapdragon also has whitish purple flowers AND the T. laxum population with the w/p flowers is the only one (out of ~15) I've found interspersed with large numbers of A. cornutum. I'll have to get MUCH better data for any hypotheses about its effect.
 |
| Antirrhinum cornutum, grown in lab, showing the pale purple/white flowers. |
I'm working now on the "genetics" (well, inheritance, but that's as close as I ever get to ATGC) of the polymorphisms (this one and selfing). Could w/p be recessive and heterozygous in more individuals (~25% under HW assumptions)? I don't think it is (entirely) developmentally induced, as in the first grow out, I only got this polymorphism from this population (I grew individuals out from 4 populations). More soon! Do let me know if you have other ideas about the system!
 |
| Heliothis phloxiphaga was a very common herbivore on T. laxum in 2014 (this plant had two - I didn't stage this), but nearly absent in 2015 - though it was still common on columbines and tarweeds. |
*I think this is confounded, as I watched two plants during each observation - nearby plants that were similar in size. Therefore, I suspect that it was more likely that a bee on the p/p plant would encounter a w/p than one from the w/p.


